The Structure and Function of Leaves Section Review 22
Identify basic common structures of plants
While individual institute species are unique, all share a mutual structure: a found body consisting of stems, roots, and leaves. They all ship water, minerals, and sugars produced through photosynthesis through the plant body in a like fashion. All plant species also respond to ecology factors, such as light, gravity, competition, temperature, and predation.
Learning Objectives
- Discuss features of constitute cells
- Identify the dissimilar tissue types and organ systems in plants
- Describe the main function and basic structure of stems
- Identify the construction and function of a typical foliage
- Identify the two types of root systems
Institute Cells

Figure i. A section of a pino embryo.
Why do plant cells look similar little rectangles? Look at Figure 1 and notice how all the cells seem to stack on each other, with no spaces in between. Might this allow the cells to form structures that can grow upright?
Organs in Plants?
Your body includes organ systems, such as the digestive system, made of individual organs, such equally the tum, liver, and pancreas, which work together to deport out a sure function (in this case, breaking down and absorbing food). These organs, in turn, are made of different kinds of tissues, which are groups of cells which work together to perform a specific job. For example, your stomach is made of musculus tissue to facilitate movement and glandular tissue to secrete enzymes for chemical breakdown of food molecules. These tissues, in plough, are made of cells specialized in shape, size, and component organelles, such as mitochondria for energy and microtubules for movement.
Plants, too, are fabricated of organs, which in plow are made of tissues. Found tissues, like ours, are constructed of specialized cells, which in turn contain specific organelles. It is these cells, tissues, and organs that conduct out the dramatic lives of plants.
Plant Cells
Establish cells resemble other eukaryotic cells in many ways. For example, they are enclosed by a plasma membrane and have a nucleus and other membrane-bound organelles. A typical establish cell is represented by the diagram in Figure 2.
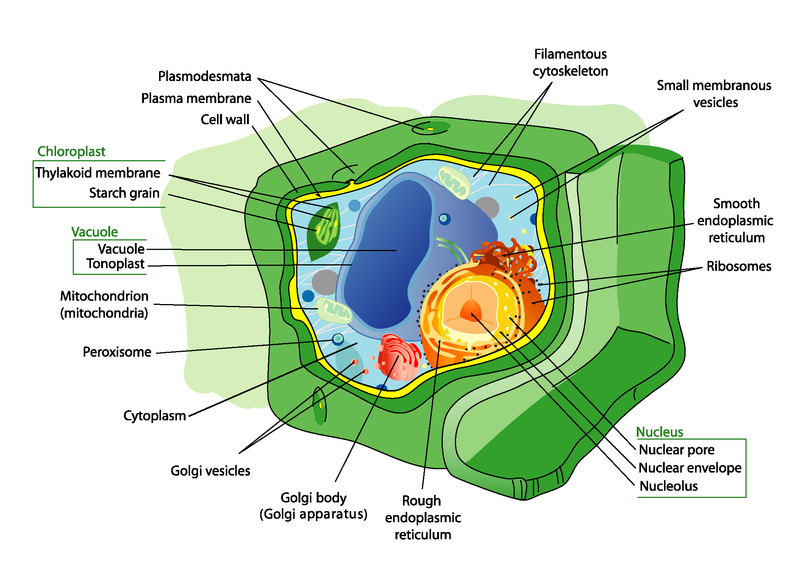
Figure 2. Plant cells have all the same structures every bit animal cells, plus some additional structures. Tin can you identify the unique plant structures in the diagram?
Plant Cell Structures
Structures found in constitute cells merely not animal cells include a large key vacuole, cell wall, and plastids such equally chloroplasts.
- The big central vacuole is surrounded by its ain membrane and contains water and dissolved substances. Its principal role is to maintain pressure against the inside of the cell wall, giving the jail cell shape and helping to support the establish.
- The cell wall is located exterior the jail cell membrane. It consists mainly of cellulose and may likewise comprise lignin , which makes information technology more than rigid. The cell wall shapes, supports, and protects the cell. It prevents the cell from absorbing too much water and bursting. Information technology too keeps large, damaging molecules out of the cell.
- Plastids are membrane-bound organelles with their own Dna. Examples are chloroplasts and chromoplasts. Chloroplasts comprise the dark-green paint chlorophyll and conduct outphotosynthesis. Chromoplasts make and store other pigments. They give flower petals their bright colors.
Types of Constitute Cells
In that location are three basic types of cells in about plants. These cells make up ground tissue, which will exist discussed in another concept. The iii types of cells are described in table below. The different types of plant cells have different structures and functions.
| Blazon of Prison cell | Structure | Functions | Example |
|---|---|---|---|
| Parenchymal | cube-shaped loosely packed thin-walled relatively unspecialized contain chloroplasts | photosynthesis cellular respiration storage | food storage tissues of potatoes |
| Collenchymal | elongated irregularly thickened walls | support current of air resistance | strings running through a stalk of celery |
| Sclerenchymal | very thick jail cell walls containing lignin | support strength | tough fibers in jute (used to brand rope) |



Plant Tissues
Plants are multicellular eukaryotes with tissue systems made of various jail cell types that carry out specific functions. Plant tissue systems autumn into i of two general types: meristematic tissue and permanent (or non-meristematic) tissue. Cells of the meristematic tissue are found in meristems, which are found regions of continuous cell partitioning and growth. Meristematic tissuecells are either undifferentiated or incompletely differentiated, and they continue to split and contribute to the growth of the plant. In contrast, permanent tissue consists of constitute cells that are no longer actively dividing.
Meristematic tissues consist of 3 types, based on their location in the institute. Apical meristems contain meristematic tissue located at the tips of stems and roots, which enable a plant to extend in length. Lateral meristems facilitate growth in thickness or girth in a maturing establish. Intercalary meristems occur but in monocots, at the bases of leaf blades and at nodes (the areas where leaves attach to a stalk). This tissue enables the monocot leaf bract to increase in length from the leaf base; for example, it allows backyard grass leaves to elongate fifty-fifty later on repeated mowing.
Meristems produce cells that chop-chop differentiate, or specialize, and go permanent tissue. Such cells take on specific roles and lose their ability to divide further. They differentiate into 3 principal types: dermal, vascular, and basis tissue. Dermal tissuecovers and protects the establish, and vascular tissue transports water, minerals, and sugars to different parts of the plant. Ground tissue serves equally a site for photosynthesis, provides a supporting matrix for the vascular tissue, and helps to store water and sugars.

Figure iii. This low-cal micrograph shows a cross section of a squash (Curcurbita maxima) stalk. Each teardrop-shaped vascular bundle consists of big xylem vessels toward the inside and smaller phloem cells toward the exterior. Xylem cells, which transport water and nutrients from the roots to the rest of the institute, are dead at functional maturity. Phloem cells, which send sugars and other organic compounds from photosynthetic tissue to the rest of the plant, are living. The vascular bundles are encased in ground tissue and surrounded by dermal tissue. (credit: modification of work by "(biophotos)"/Flickr; calibration-bar data from Matt Russell)
Secondary tissues are either elementary (composed of similar jail cell types) or complex (composed of different prison cell types). Dermal tissue, for example, is a simple tissue that covers the outer surface of the plant and controls gas exchange. Vascular tissue is an example of a circuitous tissue, and is made of two specialized conducting tissues: xylem and phloem. Xylem tissue transports water and nutrients from the roots to dissimilar parts of the found, and includes three different cell types: vessel elements and tracheids (both of which conduct h2o), and xylem parenchyma. Phloem tissue, which transports organic compounds from the site of photosynthesis to other parts of the plant, consists of four different cell types: sieve cells (which deport photosynthates), companion cells, phloem parenchyma, and phloem fibers. Unlike xylem conducting cells, phloem conducting cells are live at maturity. The xylem and phloem always prevarication side by side to each other (Figure iii). In stems, the xylem and the phloem class a construction called a vascular bundle; in roots, this is termed the vascular stele or vascular cylinder.
All animals are made of iv types of tissue: epidermal, muscle, nervus, and connective tissues. Plants, as well, are built of tissues, simply not surprisingly, their very dissimilar lifestyles derive from different kinds of tissues. All 3 types of plant cells are plant in well-nigh plant tissues. 3 major types of found tissues are dermal, footing, and vascular tissues.
Dermal Tissue
The dermal tissue of the stem consists primarily of epidermis, a single layer of cells covering and protecting the underlying tissue. Woody plants accept a tough, waterproof outer layer of cork cells normally known every bit bark, which farther protects the plant from damage. Epidermal cells are the almost numerous and least differentiated of the cells in the epidermis. The epidermis of a foliage too contains openings known as stomata, through which the exchange of gases takes place (Effigy 4). 2 cells, known every bit baby-sit cells, surround each leaf stoma, controlling its opening and closing and thus regulating the uptake of carbon dioxide and the release of oxygen and h2o vapor. Trichomes are hair-like structures on the epidermal surface. They assist to reduce transpiration (the loss of water past aboveground plant parts), increment solar reflectance, and store compounds that defend the leaves against predation by herbivores.

Figure four. Openings called stomata (singular: stoma) let a establish to take up carbon dioxide and release oxygen and water vapor. The (a) colorized scanning-electron micrograph shows a closed stoma of a dicot. Each stoma is flanked by two baby-sit cells that regulate its (b) opening and closing. The (c) baby-sit cells sit within the layer of epidermal cells (credit a: modification of work past Louisa Howard, Rippel Electron Microscope Facility, Dartmouth College; credit b: modification of work by June Kwak, Academy of Maryland; calibration-bar data from Matt Russell)
Vascular Tissue
The xylem and phloem that make upward the vascular tissue of the stem are bundled in distinct strands called vascular bundles, which run up and downwards the length of the stem. When the stem is viewed in cantankerous section, the vascular bundles of dicot stems are bundled in a ring. In plants with stems that live for more than ane twelvemonth, the individual bundles abound together and produce the characteristic growth rings. In monocot stems, the vascular bundles are randomly scattered throughout the ground tissue (Effigy 5).

Figure 5. In (a) dicot stems, vascular bundles are arranged around the periphery of the ground tissue. The xylem tissue is located toward the interior of the vascular bundle, and phloem is located toward the outside. Sclerenchyma fibers cap the vascular bundles. In (b) monocot stems, vascular bundles equanimous of xylem and phloem tissues are scattered throughout the ground tissue.
Xylem tissue has 3 types of cells: xylem parenchyma, tracheids, and vessel elements. The latter 2 types conduct water and are dead at maturity. Tracheids are xylem cells with thick secondary cell walls that are lignified. Water moves from i tracheid to another through regions on the side walls known every bit pits, where secondary walls are absent. Vessel elements are xylem cells with thinner walls; they are shorter than tracheids. Each vessel element is connected to the next by means of a perforation plate at the cease walls of the element. Water moves through the perforation plates to travel up the constitute.
Phloem tissue is composed of sieve-tube cells, companion cells, phloem parenchyma, and phloem fibers. A series of sieve-tube cells (also called sieve-tube elements) are bundled end to finish to make up a long sieve tube, which transports organic substances such as sugars and amino acids. The sugars menstruum from 1 sieve-tube jail cell to the next through perforated sieve plates, which are plant at the finish junctions between two cells. Although still live at maturity, the nucleus and other cell components of the sieve-tube cells have disintegrated. Companion cells are found alongside the sieve-tube cells, providing them with metabolic back up. The companion cells contain more ribosomes and mitochondria than the sieve-tube cells, which lack some cellular organelles.
Ground Tissue
Ground tissue is mostly made upwardly of parenchyma cells, but may also comprise collenchyma and sclerenchyma cells that help back up the stem. The basis tissue towards the interior of the vascular tissue in a stem or root is known equally pith, while the layer of tissue betwixt the vascular tissue and the epidermis is known equally the cortex.
Plant Organs
Like animals, plants incorporate cells with organelles in which specific metabolic activities have place. Unlike animals, however, plants use energy from sunlight to form sugars during photosynthesis. In addition, plant cells accept cell walls, plastids, and a large central vacuole: structures that are non found in animal cells. Each of these cellular structures plays a specific role in plant structure and function.
Lookout man Phytology Without Borders, a video produced by the Botanical Society of America about the importance of plants.
In plants, just as in animals, similar cells working together course a tissue. When dissimilar types of tissues piece of work together to perform a unique function, they form an organ; organs working together course organ systems. Vascular plants accept two distinct organ systems: a shoot organization, and a root system. The shoot arrangement consists of two portions: the vegetative (non-reproductive) parts of the found, such as the leaves and the stems, and the reproductive parts of the plant, which include flowers and fruits. The shoot system generally grows higher up basis, where it absorbs the light needed for photosynthesis. The root system, which supports the plants and absorbs water and minerals, is usually surreptitious. Figure six shows the organ systems of a typical constitute.

Figure half dozen. The shoot system of a establish consists of leaves, stems, flowers, and fruits. The root system anchors the plant while absorbing water and minerals from the soil.
Stems

Figure 7. Leaves are fastened to the plant stem at areas chosen nodes. An internode is the stem region betwixt 2 nodes. The petiole is the stem connecting the leaf to the stalk. The leaves just above the nodes arose from axillary buds.
Stems are a part of the shoot system of a plant. They may range in length from a few millimeters to hundreds of meters, and also vary in diameter, depending on the plant blazon. Stems are usually in a higher place ground, although the stems of some plants, such every bit the white potato, as well abound underground. Stems may exist herbaceous (soft) or woody in nature. Their main part is to provide support to the establish, holding leaves, flowers and buds; in some cases, stems likewise store food for the found. A stem may be unbranched, like that of a palm tree, or it may exist highly branched, similar that of a magnolia tree. The stalk of the plant connects the roots to the leaves, helping to transport absorbed water and minerals to different parts of the plant. It as well helps to transport the products of photosynthesis, namely sugars, from the leaves to the residuum of the plant.
Plant stems, whether above or below ground, are characterized by the presence of nodes and internodes (Figure seven). Nodes are points of zipper for leaves, aeriform roots, and flowers. The stem region between 2 nodes is called an internode. The stalk that extends from the stem to the base of the foliage is the petiole. An axillary bud is usually found in the axil—the expanse between the base of a leaf and the stem—where it can give rising to a branch or a flower. The apex (tip) of the shoot contains the apical meristem within the apical bud.
Stem Beefcake

Figure eight. The stem of common St John'south Wort (Hypericum perforatum) is shown in cross section in this calorie-free micrograph. (credit: Rolf-Dieter Mueller)
The stem and other plant organs arise from the ground tissue, and are primarily made upwards of simple tissues formed from 3 types of cells: parenchyma, collenchyma, and sclerenchyma cells.
Parenchyma cells are the nearly common plant cells (Figure 8). They are found in the stalk, the root, the inside of the foliage, and the pulp of the fruit. Parenchyma cells are responsible for metabolic functions, such as photosynthesis, and they help repair and heal wounds. Some parenchyma cells as well store starch. In Figure 8, nosotros come across the central pith (greenish-bluish, in the center) and peripheral cortex (narrow zone 3–5 cells thick simply within the epidermis); both are composed of parenchyma cells. Vascular tissue composed of xylem (red) and phloem tissue (green, between the xylem and cortex) surrounds the pith.
Collenchyma cells are elongated cells with unevenly thickened walls (Figure 9). They provide structural back up, mainly to the stalk and leaves. These cells are alive at maturity and are usually found below the epidermis. The "strings" of a celery stem are an example of collenchyma cells.

Figure 9. Collenchyma cell walls are uneven in thickness, as seen in this light micrograph. They provide support to institute structures. (credit: modification of piece of work by Carl Szczerski; scale-bar information from Matt Russell)
Sclerenchyma cells too provide support to the plant, merely unlike collenchyma cells, many of them are expressionless at maturity. There are two types of sclerenchyma cells: fibers and sclereids. Both types have secondary cell walls that are thickened with deposits of lignin, an organic compound that is a central component of wood. Fibers are long, slender cells; sclereids are smaller-sized. Sclereids give pears their gritty texture. Humans use sclerenchyma fibers to make linen and rope (Figure 10).

Effigy 10. The cardinal pith and outer cortex of the (a) flax stalk are made upward of parenchyma cells. Inside the cortex is a layer of sclerenchyma cells, which make upwards the fibers in flax rope and clothing. Humans have grown and harvested flax for thousands of years. In (b) this drawing, fourteenth-century women gear up linen. The (c) flax plant is grown and harvested for its fibers, which are used to weave linen, and for its seeds, which are the source of linseed oil. (credit a: modification of work by Emmanuel Boutet based on original work by Ryan R. MacKenzie; credit c: modification of work by Brian Dearth; scale-bar data from Matt Russell)
Practise Question
Which layers of the stem are fabricated of parenchyma cells?
- cortex and pith
- phloem
- sclerenchyma
- xylem
Show Reply
Reply a and b. The cortex, pith, and epidermis are fabricated of parenchyma cells.
Stem Modifications
Some plant species take modified stems that are specially suited to a detail habitat and surround (Effigy 11). A rhizome is a modified stem that grows horizontally underground and has nodes and internodes. Vertical shoots may arise from the buds on the rhizome of some plants, such as ginger and ferns. Corms are similar to rhizomes, except they are more than rounded and fleshy (such as in gladiolus). Corms contain stored food that enables some plants to survive the winter. Stolons are stems that run nearly parallel to the ground, or merely below the surface, and tin give ascent to new plants at the nodes. Runners are a blazon of stolon that runs above the ground and produces new clone plants at nodes at varying intervals: strawberries are an example. Tubers are modified stems that may store starch, as seen in the potato (Solanum sp.). Tubers arise equally swollen ends of stolons, and contain many adventitious or unusual buds (familiar to us as the "eyes" on potatoes). A bulb, which functions every bit an hugger-mugger storage unit of measurement, is a modification of a stem that has the appearance of enlarged fleshy leaves emerging from the stalk or surrounding the base of operations of the stalk, as seen in the iris.

Figure eleven. Stem modifications enable plants to thrive in a diverseness of environments. Shown are (a) ginger (Zingiber officinale) rhizomes, (b) a carrion blossom (Amorphophallus titanum) corm (c) Rhodes grass (Chloris gayana) stolons, (d) strawberry (Fragaria ananassa) runners, (e) potato (Solanum tuberosum) tubers, and (f) red onion (Allium) bulbs. (credit a: modification of piece of work past Maja Dumat; credit c: modification of work by Harry Rose; credit d: modification of work past Rebecca Siegel; credit east: modification of work by Scott Bauer, USDA ARS; credit f: modification of work by Stephen Ausmus, USDA ARS)
Watch botanist Wendy Hodgson, of Desert Botanical Garden in Phoenix, Arizona, explicate how agave plants were cultivated for food hundreds of years ago in the Arizona desert in this video: Finding the Roots of an Ancient Crop.
Some aerial modifications of stems are tendrils and thorns (Figure 12). Tendrils are slender, twining strands that enable a found (like a vine or pumpkin) to seek back up by climbing on other surfaces. Thorns are modified branches appearing as sharp outgrowths that protect the plant; common examples include roses, Osage orange and devil'south walking stick.

Figure 12. Institute in southeastern U.s., (a) buckwheat vine (Brunnichia ovata) is a weedy constitute that climbs with the assistance of tendrils. This ane is shown climbing up a wooden stake. (b) Thorns are modified branches. (credit a: modification of work by Christopher Meloche, USDA ARS; credit b: modification of work by "macrophile"/Flickr)
Leaves
Leaves are the main sites for photosynthesis: the process by which plants synthesize food. Most leaves are unremarkably green, due to the presence of chlorophyll in the foliage cells. However, some leaves may have unlike colors, caused by other institute pigments that mask the green chlorophyll.
The thickness, shape, and size of leaves are adapted to the environment. Each variation helps a constitute species maximize its chances of survival in a particular habitat. Usually, the leaves of plants growing in tropical rainforests have larger surface areas than those of plants growing in deserts or very common cold weather, which are likely to have a smaller area to minimize h2o loss.
Construction of a Typical Foliage

Figure 13. Deceptively simple in advent, a leaf is a highly efficient structure.
Each leaf typically has a leafage blade chosen the lamina, which is besides the widest part of the leaf. Some leaves are attached to the plant stem by a petiole. Leaves that exercise not have a petiole and are directly attached to the plant stalk are called sessile leaves. Small-scale green appendages usually constitute at the base of the petiole are known equally stipules. Virtually leaves have a midrib, which travels the length of the leaf and branches to each side to produce veins of vascular tissue. The edge of the leaf is called the margin. Figure thirteen shows the structure of a typical eudicot leaf.
Within each leaf, the vascular tissue forms veins. The arrangement of veins in a leaf is called the venation pattern. Monocots and dicots differ in their patterns of venation (Effigy 14). Monocots have parallel venation; the veins run in straight lines across the length of the leaf without converging at a point. In dicots, yet, the veins of the leaf accept a net-similar appearance, forming a pattern known equally reticulate venation. One extant plant, the Ginkgo biloba, has dichotomous venation where the veins fork.

Effigy 14. (a) Tulip (Tulipa), a monocot, has leaves with parallel venation. The netlike venation in this (b) linden (Tilia cordata) leaf distinguishes it as a dicot. The (c) Ginkgo biloba tree has dichotomous venation. (credit a photograph: modification of work by "Drewboy64"/Wikimedia Eatables; credit b photo: modification of work by Roger Griffith; credit c photo: modification of piece of work past "geishaboy500″/Flickr; credit abc illustrations: modification of work by Agnieszka Kwiecień)
Leaf Arrangement
The arrangement of leaves on a stem is known equally phyllotaxy. The number and placement of a plant's leaves will vary depending on the species, with each species exhibiting a characteristic leaf organization. Leaves are classified as either alternating, spiral, or opposite. Plants that accept merely one leafage per node accept leaves that are said to be either alternate—pregnant the leaves alternate on each side of the stalk in a apartment plane—or spiral, meaning the leaves are arrayed in a screw along the stem. In an opposite leaf organisation, two leaves ascend at the same betoken, with the leaves connecting reverse each other along the branch. If at that place are three or more leaves continued at a node, the leaf organisation is classified as whorled.
Leaf Class
Leaves may be simple or compound (Figure fifteen). In a simple leafage, the blade is either completely undivided—as in the banana leaf—or it has lobes, only the separation does not achieve the midrib, as in the maple leaf. In a compound leafage, the leaf blade is completely divided, forming leaflets, as in the locust tree. Each leaflet may take its ain stalk, but is attached to the rachis. A palmately chemical compound leafage resembles the palm of a hand, with leaflets radiating outwards from one signal Examples include the leaves of poison ivy, the buckeye tree, or the familiar houseplant Schefflera sp. (mutual name "umbrella plant"). Pinnately chemical compound leaves have their name from their feather-like appearance; the leaflets are bundled along the midrib, equally in rose leaves (Rosa sp.), or the leaves of hickory, pecan, ash, or walnut trees.

Figure fifteen. Leaves may be simple or compound. In simple leaves, the lamina is continuous. The (a) banana constitute (Musa sp.) has simple leaves. In compound leaves, the lamina is separated into leaflets. Compound leaves may be palmate or pinnate. In (b) palmately compound leaves, such as those of the horse chestnut (Aesculus hippocastanum), the leaflets branch from the petiole. In (c) pinnately compound leaves, the leaflets branch from the midrib, as on a scrub hickory (Carya floridana). The (d) honey locust has double chemical compound leaves, in which leaflets branch from the veins. (credit a: modification of piece of work past "BazzaDaRambler"/Flickr; credit b: modification of work by Roberto Verzo; credit c: modification of piece of work by Eric Dion; credit d: modification of work past Valerie Lykes)
Leaf Structure and Function
The outermost layer of the leaf is the epidermis; it is nowadays on both sides of the leaf and is called the upper and lower epidermis, respectively. Botanists telephone call the upper side the adaxial surface (or adaxis) and the lower side the abaxial surface (or abaxis). The epidermis helps in the regulation of gas exchange. It contains stomata (Effigy xvi): openings through which the exchange of gases takes place. Two guard cells surround each stoma, regulating its opening and closing.

Figure 16. Visualized at 500x with a scanning electron microscope, several stomata are clearly visible on (a) the surface of this sumac (Rhus glabra) leaf. At 5,000x magnification, the guard cells of (b) a unmarried stoma from lyre-leaved sand cress (Arabidopsis lyrata) have the advent of lips that environs the opening. In this (c) light micrograph cantankerous-department of an A. lyrata leaf, the guard cell pair is visible along with the large, sub-stomatal air space in the leafage. (credit: modification of piece of work by Robert R. Wise; role c scale-bar data from Matt Russell)
The epidermis is usually ane cell layer thick; nevertheless, in plants that grow in very hot or very cold conditions, the epidermis may be several layers thick to protect against excessive water loss from transpiration. A waxy layer known as the cuticle covers the leaves of all institute species. The cuticle reduces the rate of water loss from the leafage surface. Other leaves may accept small hairs (trichomes) on the leaf surface. Trichomes help to deter herbivory by restricting insect movements, or by storing toxic or bad-tasting compounds; they can also reduce the rate of transpiration by blocking air flow across the leaf surface (Effigy 17).

Figure 17. Trichomes give leaves a fuzzy advent as in this (a) sundew (Drosera sp.). Leaf trichomes include (b) branched trichomes on the leaf of Arabidopsis lyrata and (c) multibranched trichomes on a mature Quercus marilandica leaf. (credit a: John Freeland; credit b, c: modification of work by Robert R. Wise; scale-bar data from Matt Russell)
Below the epidermis of dicot leaves are layers of cells known as the mesophyll, or "centre foliage." The mesophyll of near leaves typically contains two arrangements of parenchyma cells: the palisade parenchyma and spongy parenchyma (Figure eighteen). The palisade parenchyma (also called the palisade mesophyll) has column-shaped, tightly packed cells, and may be present in one, two, or three layers. Below the palisade parenchyma are loosely arranged cells of an irregular shape. These are the cells of the spongy parenchyma (or spongy mesophyll). The air space constitute betwixt the spongy parenchyma cells allows gaseous exchange betwixt the leaf and the exterior temper through the stomata. In aquatic plants, the intercellular spaces in the spongy parenchyma help the leafage float. Both layers of the mesophyll contain many chloroplasts. Guard cells are the simply epidermal cells to incorporate chloroplasts.
In the leaf drawing (Effigy 18a), the cardinal mesophyll is sandwiched between an upper and lower epidermis. The mesophyll has ii layers: an upper palisade layer comprised of tightly packed, columnar cells, and a lower spongy layer, comprised of loosely packed, irregularly shaped cells. Stomata on the leaf underside allow gas exchange. A waxy cuticle covers all aerial surfaces of land plants to minimize water loss. These leaf layers are clearly visible in the scanning electron micrograph (Effigy 18b). The numerous small bumps in the palisade parenchyma cells are chloroplasts. Chloroplasts are likewise present in the spongy parenchyma, but are not as obvious. The bumps protruding from the lower surface of the leave are glandular trichomes, which differ in structure from the stalked trichomes in Effigy 17.
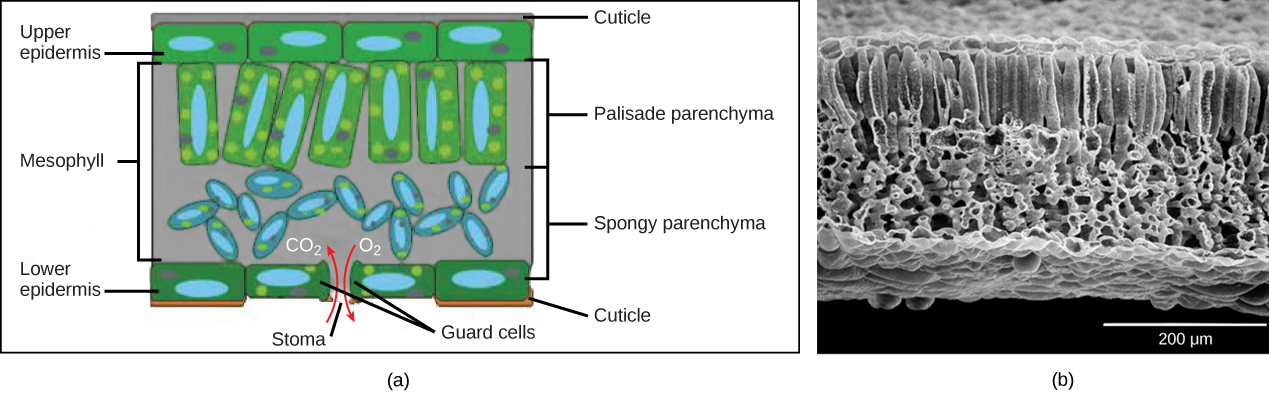
Effigy 18. (a) Foliage drawing (b) Scanning electron micrograph of a leaf. (credit b: modification of piece of work by Robert R. Wise)

Effigy 19. This scanning electron micrograph shows xylem and phloem in the leaf vascular parcel from the lyre-leaved sand cress (Arabidopsis lyrata). (credit: modification of piece of work past Robert R. Wise; calibration-bar data from Matt Russell)
Like the stem, the leaf contains vascular bundles composed of xylem and phloem (Figure 19). The xylem consists of tracheids and vessels, which transport water and minerals to the leaves. The phloem transports the photosynthetic products from the leaf to the other parts of the plant. A single vascular bundle, no matter how big or modest, always contains both xylem and phloem tissues.
Leaf Adaptations
Coniferous institute species that thrive in cold environments, similar spruce, fir, and pine, accept leaves that are reduced in size and needle-like in appearance. These needle-similar leaves take sunken stomata and a smaller surface area: two attributes that aid in reducing water loss. In hot climates, plants such as cacti have leaves that are reduced to spines, which in combination with their succulent stems, help to conserve water. Many aquatic plants take leaves with wide lamina that can float on the surface of the water, and a thick waxy cuticle on the leaf surface that repels water.
Watch "The Pale Bullpen Plant" episode of the video series Plants Are Cool, Also, a Botanical Society of America video about a carnivorous constitute species establish in Louisiana.
In Summary: Leaves
Leaves are the main site of photosynthesis. A typical leaf consists of a lamina (the broad part of the leaf, besides called the blade) and a petiole (the stalk that attaches the leaf to a stem). The system of leaves on a stem, known as phyllotaxy, enables maximum exposure to sunlight. Each constitute species has a characteristic leaf arrangement and class. The pattern of leafage organization may be alternating, opposite, or spiral, while leaf form may be uncomplicated or compound. Foliage tissue consists of the epidermis, which forms the outermost jail cell layer, and mesophyll and vascular tissue, which make up the inner portion of the leafage. In some plant species, foliage class is modified to form structures such every bit tendrils, spines, bud scales, and needles.
Roots
The roots of seed plants have three major functions: anchoring the plant to the soil, absorbing water and minerals and transporting them upwards, and storing the products of photosynthesis. Some roots are modified to absorb moisture and substitution gases. Most roots are secret. Some plants, however, also accept adventitious roots, which emerge above the ground from the shoot.
Types of Root Systems
Root systems are mainly of two types (Figure twenty). Dicots take a tap root organization, while monocots have a gristly root organisation. A tap root arrangement has a master root that grows down vertically, and from which many smaller lateral roots arise. Dandelions are a expert instance; their tap roots usually break off when trying to pull these weeds, and they can regrow another shoot from the remaining root). A tap root system penetrates deep into the soil. In dissimilarity, a fibrous root organisation is located closer to the soil surface, and forms a dumbo network of roots that as well helps prevent soil erosion (lawn grasses are a good instance, as are wheat, rice, and corn). Some plants have a combination of tap roots and gristly roots. Plants that grow in dry areas often have deep root systems, whereas plants growing in areas with arable water are likely to accept shallower root systems.

Figure twenty. (a) Tap root systems take a main root that grows down, while (b) fibrous root systems consist of many small roots. (credit b: modification of work by "Austen Squarepants"/Flickr)
Root Growth and Anatomy

Figure 21. A longitudinal view of the root reveals the zones of cell division, elongation, and maturation. Prison cell division occurs in the apical meristem.
Root growth begins with seed formation. When the plant embryo emerges from the seed, the radicle of the embryo forms the root system. The tip of the root is protected past the root cap, a structure exclusive to roots and unlike any other establish construction. The root cap is continuously replaced because it gets damaged easily as the root pushes through soil. The root tip can be divided into three zones: a zone of prison cell division, a zone of elongation, and a zone of maturation and differentiation (Figure 21). The zone of cell partitioning is closest to the root tip; it is fabricated up of the actively dividing cells of the root meristem. The zone of elongation is where the newly formed cells increase in length, thereby lengthening the root. Beginning at the outset root hair is the zone of cell maturation where the root cells begin to differentiate into special jail cell types. All three zones are in the showtime centimeter or so of the root tip.
The root has an outer layer of cells called the epidermis, which surrounds areas of ground tissue and vascular tissue. The epidermis provides protection and helps in absorption. Root hairs, which are extensions of root epidermal cells, increase the surface expanse of the root, profoundly contributing to the absorption of water and minerals.

Effigy 22. Staining reveals different cell types in this light micrograph of a wheat (Triticum) root cantankerous section. Sclerenchyma cells of the exodermis and xylem cells stain reddish, and phloem cells stain blueish. Other prison cell types stain black. The stele, or vascular tissue, is the area within endodermis (indicated past a green ring). Root hairs are visible outside the epidermis. (credit: scale-bar information from Matt Russell)
Within the root, the footing tissue forms two regions: the cortex and the pith (Figure 22). Compared to stems, roots have lots of cortex and trivial pith. Both regions include cells that shop photosynthetic products. The cortex is between the epidermis and the vascular tissue, whereas the pith lies betwixt the vascular tissue and the heart of the root.
The vascular tissue in the root is arranged in the inner portion of the root, which is chosen the stele (Effigy 23). A layer of cells known as the endodermis separates the stele from the ground tissue in the outer portion of the root. The endodermis is sectional to roots, and serves as a checkpoint for materials entering the root's vascular organisation. A waxy substance called suberin is present on the walls of the endodermal cells. This waxy region, known as the Casparian strip, forces h2o and solutes to cross the plasma membranes of endodermal cells instead of slipping betwixt the cells. This ensures that only materials required by the root laissez passer through the endodermis, while toxic substances and pathogens are more often than not excluded. The outermost jail cell layer of the root'south vascular tissue is the pericycle, an area that can give rise to lateral roots. In dicot roots, the xylem and phloem of the stele are arranged alternately in an X shape, whereas in monocot roots, the vascular tissue is bundled in a ring around the pith.

Figure 23. In (left) typical dicots, the vascular tissue forms an Ten shape in the center of the root. In (right) typical monocots, the phloem cells and the larger xylem cells grade a characteristic band around the key pith.
Root Modifications

Figure 24. Many vegetables are modified roots.
Root structures may be modified for specific purposes. For example, some roots are bulbous and shop starch. Aerial roots and prop roots are 2 forms of aboveground roots that provide additional back up to anchor the plant. Tap roots, such as carrots, turnips, and beets, are examples of roots that are modified for food storage (Figure 24).
Epiphytic roots enable a constitute to grow on another plant. For case, the epiphytic roots of orchids develop a spongy tissue to blot moisture. The banyan tree (Ficus sp.) begins as an epiphyte, germinating in the branches of a host tree; aerial roots develop from the branches and somewhen accomplish the ground, providing additional back up (Effigy 25). In screwpine (Pandanus sp.), a palm-like tree that grows in sandy tropical soils, aboveground prop roots develop from the nodes to provide additional support.

Effigy 25. The (a) banyan tree, besides known every bit the strangler fig, begins life as an epiphyte in a host tree. Aeriform roots extend to the basis and support the growing plant, which eventually strangles the host tree. The (b) screwpine develops aboveground roots that aid support the plant in sandy soils. (credit a: modification of piece of work by "psyberartist"/Flickr; credit b: modification of work by David Eikhoff)
Exercise Questions
Compare a tap root system with a fibrous root system. For each type, name a plant that provides a food in the homo diet. Which type of root system is constitute in monocots? Which type of root organisation is found in dicots?
What might happen to a root if the pericycle disappeared?
Evidence Answer
The root would not be able to produce lateral roots.
Check Your Understanding
Respond the question(s) below to run into how well you understand the topics covered in the previous section. This short quiz doesnon count toward your grade in the form, and y'all tin retake information technology an unlimited number of times.
Use this quiz to check your agreement and decide whether to (1) study the previous department further or (two) move on to the next section.
Source: https://courses.lumenlearning.com/wmopen-biology2/chapter/plant-structures/
0 Response to "The Structure and Function of Leaves Section Review 22"
Post a Comment